Eastern American Toad
Anaxyrus americanus americanus
Common Name: |
Eastern American Toad |
Scientific Name: |
Anaxyrus americanus americanus |
Etymology: |
|
Genus: |
Anaxyrus is Greek meaning "A king or chief" |
Species: |
americanus refers to America |
Subspecies: |
americanus refers to America |
Average Length: |
2 - 3.5 in. (5.1 - 9 cm) |
Virginia Record Length: |
|
Record length: |
4.4 in. (11.1 cm) |
Physical Description - This is a large toad ranging in size from 2 to 4.25 inches (50 to 107 mm). Color is variable but is usually brown, gray, olive, or brick red. There is often a light middorsal stripe. One or two warts are present in each dorsal spot. Large spiny warts also cover the dorsal part of the hind legs, particularly the tibia. This toad has well-developed cranial crests and large parotoid glands. The parotoid gland is either separate from the ridge behind the eye or connected by a short spur. The chest and anterior portion of the abdomen are usually sprinkled with dark pigment. Males are smaller than females with tubercles on the first and second fingers and have dark throats. Tadpoles range from 3/4-1 inch (1.8-2.4 cm) with body/tail length ratio 1:1.3. Body is dark and somewhat flattened. Eyes are small and dorsal. This species closely resembles the Southern Toad and the Fowler's Toad. These species frequently hybridize making identification difficult.

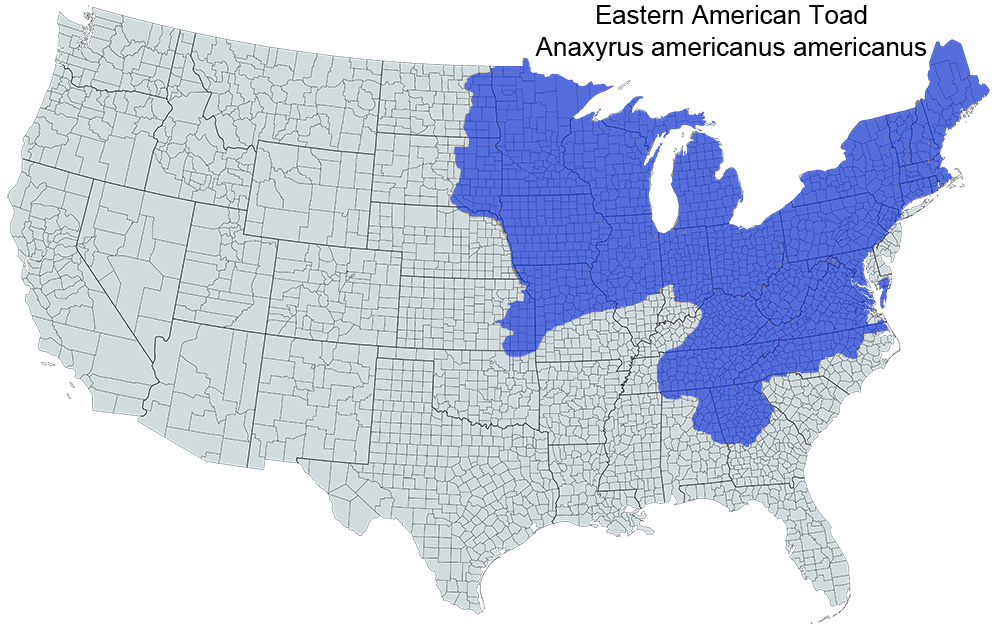
Historical versus Current Distribution - In the United States, American Toads (Anaxyrus americanus) are found throughout the eastern states from the Canadian border south to the edge of the Coastal Plain (Conant and Collins, 1998). The western edge is a line running south from western Minnesota and extreme east North Dakota, through South Dakota, Nebraska, Kansas, and Oklahoma to northeast Texas (Henrich, 1968; Collins, 1974, 1982; Seifert, 1978; Cochran, 1986a; Olson, 1987; Oldfield and Moriarty, 1994; Whiting and Price, 1994). In the south, the edge of the range runs from northeast Texas across northern Louisiana, except for an extension south along the Mississippi River to about Baton Rouge and across the middle of Mississippi (Dundee and Rossman, 1989; Lazell and Mann, 1991; Whiting and Price, 1994; Himes and Bryan, 1998). There is a gap in northwest Alabama, and the range bulges into northeast Alabama and northern Georgia (Mount, 1975). Toward the Atlantic Coast, the range encompasses only extreme northwest South Carolina, misses southeast North Carolina, and approaches the coast from Virginia and Maryland (Miller, 1979) north through New England (Hunter et al., 1992; Klemens, 1993) except for Delaware, coastal New Jersey, and extreme southeast New York including Long Island. American Toads are found throughout the Great Lakes region and northeastern states to Canada (Corin, 1976; Kraus and Schuett, 1982; Cochran, 1986b; Harding, 1997). A gap in the range occurs in southern Illinois and southwest Indiana (Garman, 1892; Smith, 1947, 1961; Minton, 1972). American Toads are found ≤ 1,200 m in the peaks of the Allegheny Mountains (Green and Pauley, 1987) and to > 1,524 m (5,000 ft) in the Great Smoky Mountains (Huheey and Stupka, 1967). Comparisons of maps in Conant and Collins (1998), Conant (1958a, 1975), and Wright and Wright (1949) reveal no fundamental changes in the species’ range during the past 50 yr. The current distribution probably reflects the distribution prior to Euro American settlement; however, many populations within this range have likely been extirpated, leaving many more populations isolated.
Two subspecies are currently recognized: Eastern American Toads (A. a. americanus) and dwarf American Toads (A. a. charlesmithi). The Dwarf American Toads is not found in Virginia. Dwarf American Toads are smaller and redder and have an unspotted belly, unlike the often profusely spotted underside of Eastern American Toads (Blair, 1943; Smith, 1961). Dwarf American Toads occupy the southwest of the species’ range, and there is a wide band of intergradation between these two subspecies; in Missouri it lies between St. Louis to an area south of Kansas City (Johnson, 1987), in Kentucky in the Jackson Purchase area (Barbour, 1971), and in Illinois through its range in the southern third of the state (Smith, 1961). A northern and colorful subspecies in Canada, A. a. copei (Ashton et al., 1973) is no longer recognized as it cannot be distinguished genetically from other populations (Guttman, 1975) despite Gaige’s (1932) morphological diagnosis. Cook (1983) proposed the synonymy of Anaxyrus americanus and Canadian toads (A. hemiophrys) based upon morphological variation across a cline between the two forms in eastern Manitoba and recognized A. a. hemiophrys as a subspecies of A. americanus. Although this system had been followed by several publications originating in Canada (e.g., Cook and Cook, 1981; Preston, 1982; Kuyt, 1991), the nomenclature advocated by Cook generally has not been accepted (Green, 1983; Frost, 1985; Collins, 1997; Conant and Collins, 1998; Green and Pustowka, 1998). American Toads were held for some time to be in taxonomic synonomy with southern toads (Anaxyrus terrestris) as Anaxyrus. t. americanus (Netting and Goin, 1946), but have been considered distinct from that species since the publication of Schmidt's (1953) checklist. Based on sequences of mitochondrial genes, Masta et al. (2002) found that American Toads are more closely related to Woodhouse's Toads (Anaxyrus woodhousii) than to Southern Toads or any other species.
Historical versus Current Abundance - Always noted as common, even "ubiquitous" (Barbour, 1971; Kolozsvary and Swihart, 1999), American Toads are frequently abundant throughout their range (Wright and Wright, 1949). Conant and Collins (1998) call American Toads the "common and abundant 'hoptoad' of the Northeast." Nowhere is this species considered to be under any threat of extirpation (Lannoo, 1998b). Nevertheless, this species is subject to fluctuations in population size (Casebere and Taylor, 1976; Christein and Taylor, 1978; Heyer, 1979; Hecnar and M’Closkey, 1996).
American Toads do not appear to be as sensitive to habitat fragmentation as are many other species of co occurring amphibians (Hager, 1998; Kolozsvary and Swihart, 1999; Lehtinen et al., 1999). They are the most readily able amphibian to re invade clearcut and burned areas of forest (Blymyer and McGinnes, 1977; Kirkland et al., 1996). However, larvae are susceptible to low pH due to acid precipitation which sublethally affects their nitrogen balance (Clark and LaZerte, 1985, 1987; Dale et al., 1985a,b; Tattersall and Wright, 1996) and thus, ultimately their survival (Clark and Hall, 1985; Freda and Dunson, 1985; Leftwich and Lilly, 1992). In consequence, acid precipitation has affected the distribution of American Toads in affected regions of their range (Clark, 1986a; Wyman, 1988a). Enhanced ultraviolet radiation, particularly UV B, however, has not been demonstrated to have a substantially detrimental effect upon American Toad eggs or larvae under natural conditions, largely due to the rapid attenuation of UV B penetration into the water (Grant and Licht, 1995; Crump et al., 1999). Chemical contaminants, including pesticides such as benzopyrene, methoxychlor, toxaphene, and endrin, as well as lead and ammonium nitrate, have both lethal and sublethal effects upon American Toads, particularly at the larval stage (Clark and Diamond, 1971; Hall and Swineford, 1979, 1981; Bracher and Bider, 1982; Steele et al., 1991, 1999; Hecnar, 1995; Jofre and Karasov, 1999; Harris et al., 2000) against which they appear to have no defense. Hindlimb deformities have been noted among juvenile toads in agricultural habitats fraught with pesticides, although to a more limited extent than among some other co occurring species, especially ranids (Ouellet et al., 1997a). Metamorphosing individuals may be particularly susceptible to certain forms of environmental stress, as they have reduced high temperature tolerance compared to both larvae and adults (Hathaway, 1928; Cupp, 1980).
Breeding migrations - American Toad males move to breeding sites, to which they have some measure of site fidelity (Ewert, 1969), and begin to call almost immediately after emergence from winter hibernation. Males call in large choruses usually beginning in early evening (Aronson, 1944a; Bogert, 1960; Brown and Littlejohn, 1972; Gerhardt, 1975; Tyning, 1990) but will sing during the daytime on particularly warm and humid days at the height of the breeding season (Dirig, 1978). In Alabama, movement to breeding sites may commence as early as mid January or February, although the peak of breeding activity is usually mid March (Mount, 1975). Toads begin to breed in late February in Louisiana (Dundee and Rossman, 1989); February–March in the Carolinas (Martof et al., 1980); early March in West Virginia (Green and Pauley, 1987); March with a peak in mid April in Kentucky (Barbour, 1971); mid March to early April in Connecticut (Klemens, 1993); late March to early April in Massachusetts (Miller, 1909b); March to early May in Kansas (Collins, 1982); mid April to mid May in Maine (Hunter et al., 1992); late April to early May in Wisconsin (Vogt, 1981) and the Great Lakes region (Werner and McCune, 1979; Harding, 1997); and early May to mid June in Minnesota (Oldfield and Moriarty, 1994). The peak of breeding activity usually lasts just under 2 wk (Hunter et al., 1992; Harding, 1997), although even after the peak, there are persistent toads that will sing even into the first days of June in more northern localities (Aronson, 1944a; Klemens, 1993).
Females appear at the breeding sites from a few days to one week after the males (Miller, 1909b; Aronson, 1944a; Collins and Wilbur, 1979) and do not persist there for as long a time. Some authors have noted an apparently non-random bias in mating based on size (Licht, 1976; Gatz, 1981a; Fairchild, 1984), but females appear to choose their mates depending upon individual characteristics of the males’ calls (Wilbur et al., 1978; Howard, 1988; Sullivan, 1992a; Howard and Palmer, 1995; Howard and Young, 1998) and orient towards the calls of particular males (Schmidt, 1971). This results in competition among males in breeding choruses for mates (Kruse, 1982; Wells and Taigen, 1984). Depending upon the density of the breeding congregation, a proportion of the males present will not sing but nevertheless attempt to intercept females as they arrive (Forester and Thompson, 1998). Males recognize toads of their own sex, regardless of species, with the aid of a well-developed and stereotypic release call (Leary, 2001b).
Breeding habitat - American Toads congregate to breed in shallow, often grassy, areas within lakes, ponds, streams, ephemeral wetlands, prairie potholes, farm ponds, floodplain pools, ditches, or streamside pools (Miller, 1909a; Wright and Wright, 1949; Mount, 1975; Dundee and Rossman, 1989; Holomuzki, 1995).
Egg deposition sites - Eggs are 1.0–2.0 mm in diameter (Wright and Wright, 1949; Smith, 1961; Dundee and Rossman, 1989) and are laid in two long strings, usually entwined around vegetation or resting on the pond bottom (Miller, 1909a; Wright and Wright, 1949; Tyning, 1990; Holomuzki, 1997; Behler and King, 1998) in shallow water 5–10 cm deep (Miller, 1909a). ii. Clutch size. Clutch size has been reported to range from 2,000–12,000 eggs (Gilhen, 1984); 4,000–12,000 eggs (Livezey and Wright, 1947); 4,000–8,000 eggs (Wright and Wright, 1949); 4,000–15,000 eggs (Miller, 1909a); or 2,000–20,000 eggs (Johnson, 1987). Numbers of eggs/in (2.5 mm) vary from 15–17 in eastern American Toads to a mean of about 7.5/in dwarf American Toads (Livezey and Wright, 1947; Dundee and Rossman, 1989).
Altig & McDiarmid 2015 - Classification and Description:
- Eastern Linear
- Arrangement 3 - Eggs oviposited as strings.
- Sub-arrangement C - Jelly tube bilayered and obvious, string long. Eggs deposited in nonflowing water and lake margins; Ovum Diameter 1.0-1.4 mm; tube diameter 3.4-4.0 mm; partitions present between uniserial ova.
- Arrangement 3 - Eggs oviposited as strings.
Length of larval stage - Eggs hatch in 3–12 d (Smith, 1961; Martof et al., 1980; Vogt, 1981). The larval period takes 50–60 d (Wright and Wright, 1949), and tadpoles transform at 7–12 mm SVL (Wright and Wright, 1949) or 6–8 mm SVL in dwarf American Toads (Dundee and Rossman, 1989). High tadpole density negatively affects growth and timing of metamorphosis (Brockelman, 1969; Wilbur, 1977a; Breden and Kelly, 1982).
Larval requirements - American Toad tadpoles are black and reach a length of about 2.5 cm (Altig, 1970; Dundee and Rossman, 1989; Oldfield and Moriarty, 1994). They evidently require small, open, still water ponds and pools devoid of fishes (Wright and Wright, 1949), including ephemeral ponds (Wilbur, 1990), preferentially not overly shaded by trees (Werner and Glennemeier, 1999).
Tadpole food - Tadpoles are omnivorous and will consume vegetable matter, suspended organic matter and algae, detritus, and dead fish or tadpoles (Test and McCann, 1976; Seale and Beckvar, 1980; Bowen, 1984; Ahlgren and Bowen, 1991). American Toad tadpoles compete for food with the tadpoles of other anuran species to variable extents (Alford and Wilbur, 1985; Wilbur and Alford, 1985; Alford, 1989a,b).
Tadpoles:
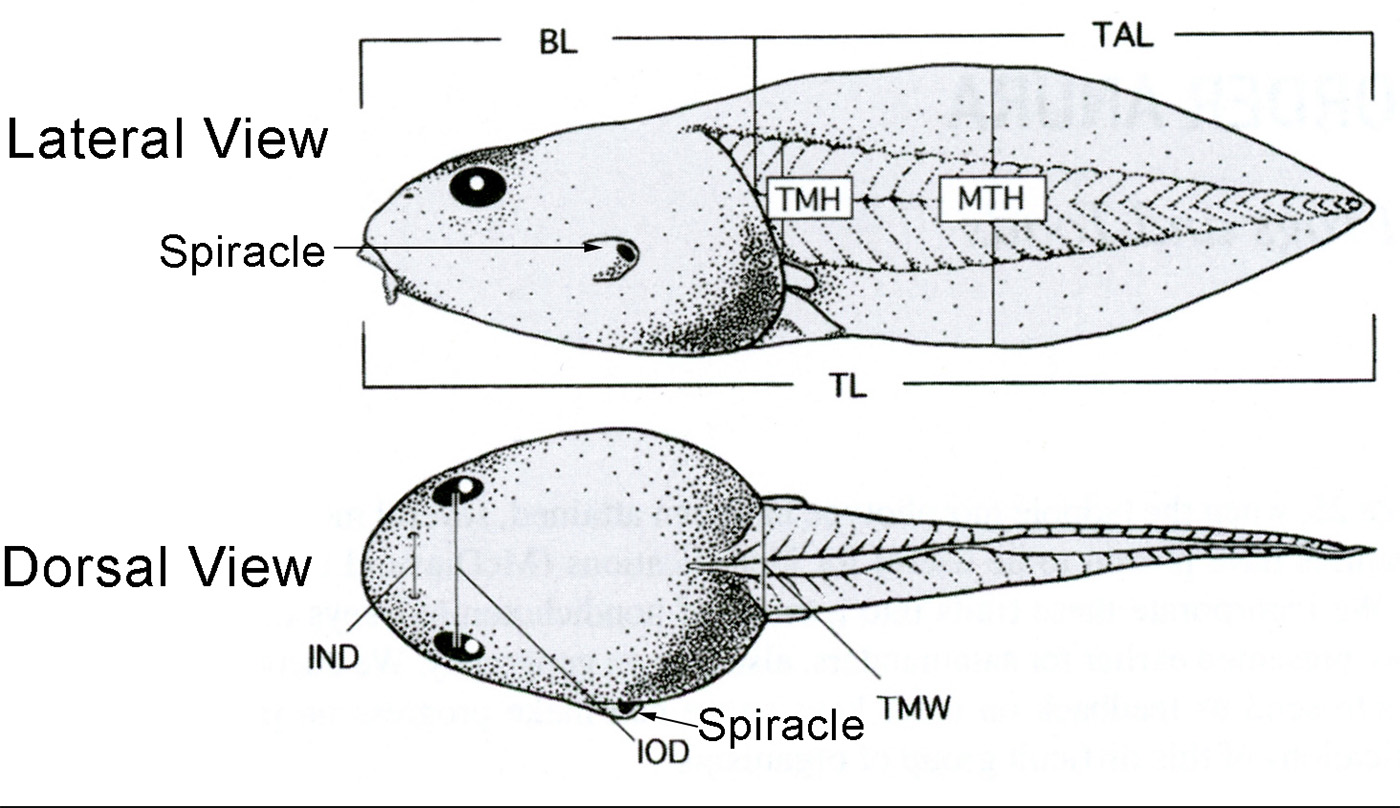
| Lateral View | Dorsal View |
|---|---|
| BL = Body Length | IND = Internarial Distance |
| MTH = Maximum Tail Height | IOD = Interorbital Distance |
| TAL = Tail Length | TMW = Tail Muscle Width |
| TL = Total Length | |
| TMH = Tail Muscle Height |
Features of metamorphosis - Metamorphosis occurs in mid May in Louisiana (Dundee and Rossman, 1989) and West Virginia (Green and Pauley, 1987); early June in Illinois (Smith, 1961); mid June in Massachusetts (Miller, 1909a); mid June to early August in Connecticut (Klemens, 1993); late June to early July in Wisconsin (Vogt, 1981); and late July in Minnesota (Oldfield and Moriarty, 1994). Metamorphosis is rapid (Miller, 1909a); newly metamorphosed animals will quit the water within 1 d of acquiring their front legs and will have completely resorbed the tail a mere 2 d later. A whole cohort will transform within 6 d.
Post-metamorphic migrations - Newly metamorphosed animals and juveniles disperse widely but not aimlessly, evidently orienting themselves in part through use of celestial cues (Dole, 1972c, 1973).
Juvenile Habitat - Similar to adults, although large aggregations of post-metamorphic toads can often been seen on mudflats surrounding breeding wetlands. American Toads grow rapidly following transformation, doubling in length and more than quadrupling their weight from June–August (Hamilton, 1934).
Adult Habitat - American Toads evidently tolerate humans well. They can be common in gardens, fields, lawns, and barnyards (Dickerson, 1906; Smith, 1947, 1961; Klemens, 1993). Mainly nocturnal, during the day toads seek cover under stones, boards, woodpiles, walkways, porches, or other cover (Wright and Wright, 1949). American Toads are tolerant of brackish estuarine waters (Kiviat and Stapleton, 1983). Mount (1975), in Alabama, and Johnson (1987), in Missouri, note that American Toads are common in and along the edges of forested areas. Oldfield and Moriarty (1994) note that American Toads will thrive in prairie parkland habitats, although they seem to be predominantly animals of the forest, a point contradicted by Guerry and Hunter, who found a negative association of American Toads in relation to forest area. Green and Pauley (1987) observed that in West Virginia individuals may be found in dense woods, but are more frequently found in more open pastures, agricultural areas, and gardens. Fitch (1958) considered the optimum habitat for American Toads in Kansas to be rocky habitats in open woods or wooded edges, where toads were found under large flat rocks covering loose, damp soil. Nevertheless, due to their predilection for orchards, gardens, and lawns in Illinois, Smith (1947) considered American Toads to be essentially a prairie species.
Home Range Size - Fitch (1958) and Collins (1974, 1982) report that American Toads in Kansas will establish home ranges of 0.16 ac or more in size. Toads will repeatedly use particular hiding places (Dole, 1972c) and will return daily to "forms," a small depression in a lawn or under stones (M. Stewart, personal observations), but during wet periods will travel great distances ≤ 1 km (Ewert, 1969).
Territories - Not territorial (Fairchild, 1984; Tyning, 1990).
Aestivation/Avoiding Dessication - In summer, American Toads may be little in evidence during extended periods of hot, dry weather. Toads likely aestivate during the summer in Alabama (Mount, 1975).
Seasonal Migrations - Toads move in spring to breeding sites from wherever they had been hibernating during the winter (Miller, 1909a; Oldham, 1969), both over land and along streams (Maynard, 1934). Weather, particularly humidity and rainfall, influences movement (Fitzgerald and Bider, 1974a,b).
Nocturnal postbreeding movements have been recorded ≤ 1,000 m, but most excursions are considerably shorter (Ewert, 1969). Dole (1972c) found that toads displaced ≤ 235 m from point of capture will move and orient themselves but not necessarily towards the point of capture. Both vision and olfaction are used by adult toads to orient (Dole, 1972c; Adler and Taylor, 1981).
Torpor (Hibernation) - During cold winter weather, American Toads hibernate terrestrially (Miller, 1909b). They dig backwards into the soil where they reside or find another hibernation site that permits them to burrow below the frostline (Wright and Wright, 1949; Tester et al., 1965; Ewert, 1969). American Toads are not freeze tolerant (Miller, 1909b; Storey and Storey, 1986) and evidently have no mechanism for freeze tolerance (Holzwart and Hall, 1984). Hibernation begins as the temperature falls below their normal activity minimum of about 9 ˚C, which is usually October in northern U.S. populations (Oldfield and Moriarty, 1994).
Interspecific Associations/Exclusions - The members of the Anaxyrus americanus group of toads (W.F. Blair, 1963a, 1972a; Guttman, 1969; Martin, 1973) are notorious hybridizers (A.P. Blair, 1941; Sanders, 1961, 1987; W.F. Blair, 1964a, 1972b; Green, 1983, 1984, 1996; Green and Parent, 2003). American Toads form a hybrid zone in the northwest with Canadian toads (Henrich, 1968, Cook, 1983, Green, 1983, Green and Pustowka, 1997) and another in the southeast with southern toads (Neill, 1949a; Volpe, 1955b; Mount, 1975; Weatherby, 1982). In Louisiana and Alabama, American Toads are not usually found breeding in sympatry with southern toads, due to some separations in range and because American Toads breed earlier (Mount, 1975; Dundee and Rossman, 1989) by 4–6 wk. The same difference in timing of breeding is also true for American Toads relative to Canadian toads and Fowler’s toads (B. fowleri); American Toads breed earlier and at colder temperatures (Henrich, 1968; Barbour, 1971; Green, 1982). Nevertheless, overlaps in breeding seasons do occur (Green, 1984). American Toads are broadly sympatric with Fowler’s toads, with which they are known to hybridize in numerous scattered localities (Allard, 1908; Miller and Chaplin, 1910; Hubbs, 1918; Myers, 1927; Pickens, 1927a; Blair, 1941; Volpe, 1952, 1955b; Cory and Manion, 1955; Zweifel, 1968a; L.E. Brown, 1970; Jones, 1973; Green, 1982, 1984; Green and Parent, 2003). The two species tend to have different temperature tolerances (Frost and Martin, 1971) and habitat preferences: American Toads in forests, Fowler’s toads in more open sandy areas and savannas (Smith, 1961; Mount, 1975; Green and Pauley, 1987; Johnson, 1987). There had been long-standing confusion between the two species in the literature until their identities were clearly defined by Myers (1927, 1931b) and explained by Wright and Wright (1949). Dickerson (1906), for instance, includes a photograph labeled American Toads although the animals portrayed are clearly Fowler’s toads.
Age/Size at Reproductive Maturity - American Toads are strongly sexually dimorphic in body size (Licht, 1976; Gatz, 1981a; Kruse, 1981) with an apparent south to north gradient of increasing average size. Throughout most of their range, male eastern American Toads reach 54–85 mm, but females are 56–111 mm (Wright and Wright, 1949; Conant and Collins, 1998). In Connecticut, Klemens (1993) found 33 males measuring from 51–72 mm, averaging 60.9 mm, whereas 18 females measured from 68–85 mm, averaging 75.1 mm. Particularly large individuals, ≤ 155 mm, have been reported on islands in northern Lake Michigan (Long, 1982). Dwarf American Toads are comparatively smaller, reported to be 44–70 mm in Louisiana (Dundee and Rossman, 1989). In Illinois, Smith (1961) reports that 47 eastern American Toads ranged from 47.0–90.0 mm SVL, whereas 19 dwarf American Toads ranged from 42.8–69.5 mm, stating also that dwarf American Toads are usually under 60 mm. American Toads reach sexual maturity in 2–4 yr (Dickerson, 1906; Hamilton, 1934), but the sexes differ: males are mature by the end of their second summer and breed the following spring. Females mature at the age of 3 (Acker et al., 1986; Green and Pauley, 1987) or 4 (Kalb and Zug, 1990).
Longevity - Dickerson (1906) mentions, as "authentic record," the story of an American Toad living to be 36 yr of age, but this is undoubtedly an exaggeration. Both Acker et al. (1986), in Illinois, and Kalb and Zug (1990), in Virginia, found breeding males up to 4 yr old and breeding females up to 5 yr old using skeletochronology. An American Toad of unrecorded sex survived 4 yr, 8 mo, and 25 d at the Philadelphia Zoo (Bowler, 1977), and a captive male from Illinois lived 5 yr, 6 mo and 21 d (Morris and Meyer, 1980).
Feeding Behavior - American Toads opportunistically eat large numbers of insects and other invertebrates (Miller, 1909a; Hamilton, 1934; Wright and Wright, 1949; Oliver, 1955a; Leclair and Vallieres, 1981; Gilhen, 1984; Garrett and Barker, 1987; Green and Pauley, 1987; Jennings et al., 1991; Lannoo, 1996) and their methods are lovingly described by Dickerson (1906). They eat beetles in large numbers (Dickerson, 1906; Miller, 1909a; Wright and Wright, 1949; Oliver, 1955a; Gould and Massey, 1984), with the consumption of ground beetles (Carabidae) especially noted (Larochelle, 1974, 1975a,b, 1976, 1977a,b,c; Charlebois, 1977), although they are evidently stymied by bombardier beetles (Dean, 1980a,b). They also consume large numbers of ants (Oliver, 1955a; Bellocq et al., 2000) and moths (Babbitt, 1937). Primarily crepuscular, American Toads will feed at night on insects attracted to lawn and garden lights (Klemens, 1993; Lannoo, 1996), although they will also search for food during late afternoons.
Predators - American Toads are eaten readily by eastern hog-nosed snakes (Heterodon platirhinos; Barbour, 1973), water snakes (Nerodia sp.), garter snakes (Thamnophis sp.), and occasionally by ducks (Mueller, 1980), crows, and screech owls (Miller, 1909b). Raccoons (Procyon lotor) will consume toads, generally by eating the belly and leaving the dorsal skin behind (Schaaf and Garton, 1970), as will striped skunks (Mephitis mephitis; Groves, 1980). Huheey and Stupka (1967) mention the remains of an American Toad in the stomach of a road killed opossum. American Bullfrogs (Lithobates catesbeianus) try but fail to eat toads (Tucker and Sullivan, 1975). According to Miller (1909b), "Boys are very destructive to toads in the spawning season."
American Toad tadpoles are consumed in general order of importance (Miller, 1909b) by larvae of predaceous diving beetles (Dytiscidae; Kruse, 1983; Leclair et al., 1986), newts (Notophthalmus sp.; Miller, 1909b), dragonfly naiads (Van Buskirk, 1988), giant water bugs (Belostomatidae; Miller, 1909b), and crayfish (Miller, 1909b). Least sandpipers have been known to consume them as well (Stangel, 1983).
Anti-Predator Mechanisms When confronted by a potential predator, a toad will inflate its lungs, lower its head, and lift up its body, while remaining immobile (Brown and Thrall, 1974; Dodd and Cupp, 1978; Marchisin and Anderson, 1978; Vogt, 1981). Otherwise a toad relies upon immobility and its camouflaging resemblance to a clod of dirt for protection (Heinen, 1985, 1994). The skin secretions of the warts are harmless to human skin but will irritate mucous membranes thus making the toads distasteful to many animals, including shrews (Formanowicz and Brodie, 1982) and domestic dogs (Dickerson, 1906; Green and Pauley, 1987). Toads have little defense, though, against boys (Miller, 1909b).
As in other species of Anaxyrus, toxins contained in the skin secretions are also secondarily deposited in the ova (Licht, 1968) and are thought thereby to afford some protection to newly laid, fertilized eggs, which then decreases as the zygotes develop (Phisalix, 1922).
Many vertebrate predators find the tadpoles distasteful (Brodie et al., 1978; Brodie and Formanowicz, 1987). However, invertebrate predators, as they suck body contents and do not consume the skin (Wassersug, 1973), generally are reported to find bufonid larvae palatable (Kruse, 1983; Leclair et al., 1986; Van Buskirk, 1988), even though they are repulsed by the skin secretions of newly metamorphosed animals (Brodie et al., 1978; Formanowicz and Brodie, 1982). Tadpoles will adjust their behavior depending upon the particular predator encountered (Reylea, 2001). Tadpoles respond to the presence of predatory odonate larvae, which they detect by olfaction, by decreasing feeding rate and exhibiting immobility and avoidance (Anholt et al., 1996; Petranka and Hayes, 1998). They also transform earlier at a smaller size (Skelly and Werner, 1990). Density of predators and food availability do not appear to appreciably affect these responses (Petranka, 1989a; Pearman, 1995; Anholt et al., 1996), nor does learning play a role (Gallie et al., 2001). Tadpoles commonly accrue injuries to the tail tip due to attempted predation (Blair and Wassersug, 2000). American Toad tadpoles form schools (Black, 1971a, 1975; Beiswenger, 1975, 1977, 1978; Feder, 1984; Dupre and Petranka, 1985) abetted by kin recognition (Waldman and Adler, 1979; Waldman, 1981, 1982a, 1985a,b, 1986; Dawson, 1982). Kruse and Stone (1984) found that largemouth bass (Micropterus salmoides) learned to distinguish American Toad tadpoles from spring peeper (Pseudacris crucifer) tadpoles, but surmised that this was likely due to the schooling behavior of the tadpoles that may thus also serve as a deterrent to predation. Newly metamorphosed toadlets also form aggregations that may be a part of an anti-predation defense against snakes (Hayes, 1989; Heinen, 1993a,b, 1994, 1995). Juveniles may avoid snakes by relying upon chemical clues of the snakes’ presence (Flowers and Graves, 1997). As with adults, newly metamorphosed toadlets respond to the touch of garter snakes by crouching and remaining immobile, which affords them some success in thwarting predation; unlike adults, juveniles do not noticeably inflate themselves (Hayes, 1989).
Diseases - An incidence of a xanthoma cancer has been noted by Counts and Taylor (1977), and the incidence of assorted hindlimb deformities was noted by Ouellet et al. (1997a). R. Parasites. American Toads harbor a host of internal and external parasites, including protozoans, helminths of various sorts, and other organisms. Protozoan parasites include the sarcomastigote cloacal parasite Opalina (Delvinquier and Desser, 1996), the myxosporean Myxidium serotinum (McAllister and Trauth, 1995), and the protozoan pathogen Toxoplasma (Stone and Manwell, 1969). Trypanosomiasis blood parasitism is widespread (Werner and Walewski, 1976; Barta and Desser, 1984; Werner et al., 1988; Lun and Desser, 1996), particularly involving the species Trypanosoma fallisi (Martin and Desser, 1990, 1991a,b; Martin et al., 1992, 2002). Nematode parasites include Gyrinicola batrachiensis of tadpoles (Adamson, 1981a,b) and Oswaldocruzia pipiens (Baker, 1977), Rhabdias spp. (Baker, 1978a, 1979), Cosmocercoides dukae (Baker, 1978b), and Cosmocercoides variabilis (Vanderburgh and Anderson, 1987a,b; Joy and Bunten, 1997) of adults. The apicomplexan Hepatozoon clamatae (Kim et al., 1998), trematodes of the genus Allassostomoides (Brooks, 1975), cestodes of the genus Mesocestoides, assorted other helminths (James, 1969; Ulmer, 1970; Ulmer and James, 1976; Ashton and Rabalais, 1978; D.D. Williams, 1978; Williams and Taft, 1980; Coggins and Sajdak, 1982; Bolek and Coggins, 2000), dipteran flies (Bufolucilia elongata; Briggs, 1975), the leech Desserobdella picta (Briggler et al., 2001), and the parasitic fungus Dermosporidium penneri (Jay and Pohley, 1981) are also known to parasitize American Toads.
Conservation - Conant and Collins (1998) call American Toads the "common and abundant 'hoptoad' of the Northeast." Nowhere is this species considered to be under any threat of extirpation (Lannoo, 1998b). Nevertheless, this species is subject to fluctuations in population size (Casebere and Taylor, 1976; Christein and Taylor, 1978; Heyer, 1979; Hecnar and M’Closkey, 1996).
American Toads do not appear to be as sensitive to habitat fragmentation as are many other species of co-occurring amphibians (Hager, 1998; Kolozsvary and Swihart, 1999; Lehtinen et al., 1999). They are the most readily able amphibian to re-invade clearcut and burned areas of forest (Blymyer and McGinnes, 1977; Kirkland et al., 1996). However, American Toad larvae are susceptible to low pH due to acid precipitation, which sublethally affects their nitrogen balance (Clark and LaZerte, 1985, 1987; Dale et al., 1985a,b; Tattersall and Wright, 1996) and thus, ultimately, their survival (Clark and Hall, 1985; Freda and Dunson, 1985; Leftwich and Lilly, 1992). In consequence, acid precipitation has reduced the distribution of American Toads in affected regions of their range (Clark, 1986; Wyman, 1988a). Enhanced ultraviolet radiation, particularly UV-B, has however not been demonstrated to have a substantial detrimental effect upon American Toad eggs or larvae under natural conditions, largely due to the rapid attenuation of UV-B penetration into the water (Grant and Licht, 1995; Crump et al., 1999). Chemical contaminants, including pesticides such as benzopyrene, methoxychlor, toxaphene and endrin, as well as lead and ammonium nitrate, have both lethal and sublethal effects upon American Toads, particularly at the larval stage (Clark and Diamond, 1971; Hall and Swineford, 1979, 1981; Bracher and Bider, 1982; Steele et al., 1991, 1999; Hecnar, 1995; Jofre and Karasov, 1999; Harris et al., 2000), against which they appear to have no defense. However, Allran and Karasov (2001) indicate that atrazine concentrations in the field may not be high enough to cause direct effects. Hindlimb deformities have been noted among juvenile toads in agricultural habitats fraught with pesticides, although to a more limited extent than among some other co-occurring species, especially ranids (Ouellet et al., 1997). Metamorphosing individuals may be particularly susceptible to certain forms of environmental stress, as they have reduced high temperature tolerance compared to both larvae and adults (Hathaway, 1928; Cupp, 1980).
References for Life History
- Altig, Ronald & McDiarmid, Roy W. 2015. Handbook of Larval Amphibians of the United States and Canada. Cornell University Press, Ithaca, NY. 341 pages.
- AmphibiaWeb. 2020. University of California, Berkeley, CA, USA.
- Conant, Roger and, Collins, John T., 2016, Peterson Field Guide: Reptiles and Amphibians, Eastern and Central North America, 494 pgs., Houghton Mifflin Company., New York
- Duellman, William E. and, Trueb, Linda, 1986, Biology of Amphibians, 671 pgs., The Johns Hopkins University Press, Baltimore
- Martof, B.S., Palmer, W.M., Bailey, J.R., Harrison, III J.R., 1980, Amphibians and Reptiles of the Carolinas and Virginia, 264 pgs., UNC Press, Chapel Hill, NC
- Wilson, L.A., 1995, Land manager's guide to the amphibians and reptiles of the South, 360 pp. pgs., The Nature Conservancy, Southeastern Region, Chapel Hill, NC
Photos:
*Click on a thumbnail for a larger version.









Verified County/City Occurrence
Accomack
Albemarle
Alleghany
Amelia
Amherst
Appomattox
Arlington
Augusta
Bath
Bedford
Bland
Botetourt
Brunswick
Buchanan
Buckingham
Campbell
Caroline
Carroll
Charles City
Charlotte
Chesterfield
Clarke
Craig
Culpeper
Cumberland
Dickenson
Dinwiddie
Essex
Fairfax
Fauquier
Floyd
Fluvanna
Franklin
Frederick
Giles
Gloucester
Goochland
Grayson
Greene
Greensville
Halifax
Hanover
Henrico
Henry
Highland
Isle of Wight
James City
King and Queen
King George
King William
Lancaster
Lee
Loudoun
Louisa
Lunenburg
Madison
Mathews
Mecklenburg
Montgomery
Nelson
New Kent
Northumberland
Nottoway
Orange
Page
Patrick
Pittsylvania
Powhatan
Prince Edward
Prince George
Prince William
Pulaski
Rappahannock
Richmond
Roanoke
Rockbridge
Rockingham
Russell
Scott
Shenandoah
Smyth
Southampton
Spotsylvania
Stafford
Surry
Sussex
Warren
Washington
Westmoreland
Wise
Wythe
York
CITIES
Alexandria
Chesapeake
Danville
Fairfax
Falls Church
Lynchburg
Newport News
Richmond
Roanoke
Suffolk
Verified in 92 counties and 10 cities.
U.S. Range

Box Turtle Reporting
